
A continuación, presento una extensa entrevista con el biólogo Daniel Heredia Doval sobre biología evolutiva, que espero sea del provecho de todos los estudiosos, estudiantes, investigadores y público general interesados en la biología, la evolución y las ciencias de la vida. Agradezco a Daniel su gentileza en responder detalladamente todas las preguntas planteadas. Espero que los lectores disfrunten la entrevista.
1-Daniel, ¿puedes decirnos un poco sobre tu formación académica y
qué te motivó a interesarte por el estudio científico de la naturaleza y los
seres vivos?
Soy licenciado en Biología por la Universidad Autónoma de Madrid.
En 2009 comencé mis estudios de doctorado en dicha universidad bajo la
dirección del Dr. Máximo Sandín. En la actualidad me encuentro finalizando mi
tesis doctoral, en la que argumento sobre la necesidad de un cambio de
paradigma en Biología y en la que propongo nuevos enfoques y herramientas
(categóricas, técnicas y metodológicas) que surgen de la integración de la
teoría de sistemas y las ciencias de la complejidad con las ciencias
biológicas, en una síntesis que pretende unificar los principales
planteamientos heterodoxos para responder a las principales carencias del
neodarwinismo, y que coloco bajo la etiqueta de Biología Evolutiva de Sistemas.
También soy coordinador de BiVOS, Grupo de Estudios Teóricos en
Biología Evolutiva de Sistemas, una plataforma formativa y divulgativa formada
recientemente en torno a este área emergente. Toda la información referente a
los objetivos y las actividades de BiVOS está en su página web, así como
noticias y ensayos relevantes.
Respecto a mi interés por el estudio de la naturaleza, este me
viene desde niño. Con solo trece años ya tenía claro qué quería estudiar en un
futuro, y me pasaba las tardes entre libros de divulgación y documentales de naturaleza.
Podría decir que es un interés vocacional, aunque reconozco que este
sentimiento a venido reforzado por la buena influencia de familiares,
compañeros y profesores con la misma pasión.
2-Para entrar en materia, quizás cierta precisión terminológica
sería apropiada. En los libros sobre biología, he encontrado que el término
"evolución" se emplea para referirse a muchos conceptos, entre los
cuáles se encuentran "descendencia con modificación",
"especiación", "adaptación", "cambio en la frecuencia
de los fenotipos prevalecientes en una población", "tesis del
ancestro común", "cambios de organización" etc., y tal
diversidad tiende a generar una fuerte confusión terminológica y conceptual.
¿Podrías aclararnos que significan cada uno de estos conceptos y a cuál de
ellos se refiere el término "evolución"?
Si, tal como apuntas, uno de los más graves lastres de la Biología
Evolutiva es la falta de una definición clara y concisa de muchas de las
categorías con las que se trabaja,
incluyendo los sujetos y objetos de estudio, así como una distinción
concisa entre los mecanismos, los procesos, los fenómenos y las dinámicas que
entrarían dentro del estudio de la evolución.
Los términos incluidos en el apartado hacen clara referencia al
aparato conceptual de la síntesis neodarwinista, aún sin aparecer de forma
explícita el concepto de selección natural. La descendencia con modificación es
una condición necesaria en la evolución (llamémosla creatividad
intergeneracional), pero no definitoria, dado que los procesos evolutivos
implican una homogenización poblacional que establezca una dirección, y esto no
queda descrito en la sentencia anterior. Por supuesto, el punto de vista
convencional es aplicar a esta “descendencia con modificación” un “agente
selectivo” que determine la dirección de los cambios, aunque esta no es la
única forma (ni a mi juicio la más coherente) de explicar la homogeneización de
los cambios en las poblaciones.
Los términos “especiación” y “adaptación” hacen referencia a dos
cuestiones clásicas en Biología Evolutiva. La especiación es la formación de
una nueva especie a partir de otra precursora. Hasta aquí todo es bastante
claro. Lo realmente problemático es definir con pulcritud qué es (si es que es
algo) una especie y cuando una población adquiere esta categoría taxonómica,
dado que existen varios criterios y abundantes excepciones. Respecto a la
“adaptación”, yo la definiría como el fenómeno inherente de coherencia
ecológica entre los sistemas biológicos en, y con, su medio. Por supuesto existen
otras definiciones “oficiales” que involucran otros conceptos y que se centran
en la composición poblacional (“cambio en la frecuencia de los
fenotipos/genotipos prevalecientes”), pero de nuevo, estas suelen hacer uso del
aparato neodarwinista (eficiencia biológica, reproducción diferencial, etc.)
que yo personalmente excluiría por las razones que expongo más adelante.
La “tesis del ancestro común” no es sinónimo de evolución. Es una
de las múltiples (y no siempre coherentes entre sí) propuestas de Darwin en su
obra “El origen de las especies”, y por lo tanto no es dominio del hecho
evolutivo sino de la teoría darwinista de la evolución. La idea de un ancestro
común es coherente dentro de un esquema de filogenia arborescente como el
planteado por el darwinismo, pero no necesariamente cierta. De hecho algunos
autores, como Carl Woese, no hablan de un último ancestro común, sino de una
población inicial (progenote) marcada por el consenso genético y unas altas
tasas de transferencia horizontal, siendo esta una alternativa coherente y
perfectamente plausible en vistas de la argumentación ofrecida.
Respecto a los “cambios de organización” nos encontramos con un
problema similar al de la “especiación”. Este criterio, aun siendo muy
interesante, carece de sentido en ausencia de una definición clara y concisa de
qué es una “organización” en la biología moderna. Lo más explícito a citar
serían los clásicos bauplan (planes de organización) de los principales
grupos de animales, si bien esta referencia sigue sin dar una definición
concreta y universal. De nuevo, volvemos a encontrarnos en un gran vacío
categórico. En mi opinión, es urgentemente necesario definir correctamente este
y otros términos para poder operar con una definición correcta de “Evolución” y
abarcar nuevas direcciones en biología evolutiva.
Aunque puedo dar una definición “oficial” o “extraoficial” para
cada uno de estos términos, esto no quiere decir que todos ellos sean adecuados
para incluirse a su vez en la definición del término “Evolución”. Considero que éste debería hacer referencia
únicamente al “fenómeno de cambio en la organización de los sistemas biológicos
en el tiempo, a través de mecanismos naturales”, sin una referencia directa a
las teorías e hipótesis que tratan de explicar dicho fenómeno o los mecanismos
implicados. Eso sí, lo que es ineludible es formular claramente todos y cada
uno de los elementos de esta definición, lo cual ya se saldría de esta
entrevista.
3-¿Qué se entiende por darwinismo (o neo-darwinismo) y teoría sintética
de la evolución?
Darwinismo es una corriente evolucionista cuyo origen se atribuye
a Charles Darwin, quién publicó sus ideas acerca de la variación y
“supervivencia de las razas más favorecidas en la lucha de la existencia” en su
entonces polémica obra “El origen de las especies” (1859) [1]. La idea
fundamental del darwinismo, su núcleo teórico, es que la competencia por los
recursos limitados lleva a una situación mantenida de supervivencia y
reproducción diferencial dentro de las poblaciones, de manera que los
individuos más competentes y mejor adaptados dejan descendencia mientras el
resto pasa a ser parte de la historia. Dado que las características ventajosas
de estos individuos “virtuosos” se heredan, su descendencia vuelve a obtener
una ventaja en las mismas condiciones, lo cual lleva finalmente a un remplazo
de la población y a la homogeneización de estos caracteres. En esencia de lo
que se trata es de una quimera del pensamiento poblacional de Malthus y del
liberalismo de Adam Smith “aplicada al conjunto de los reinos animal y
vegetal”. Esta idea de “supervivencia de
los más aptos” es lo que se ha denominado sin demasiada formalidad (como el
propio Darwin reconocía) como “Selección Natural”. Es importante señalar, no
obstante, que el darwinismo clásico contenía otros elementos fundamentales e
indisolubles para Darwin: gradualismo, uniformismo, adaptacionimo, ancestro
final único, variabilidad individual no finita y filogenia arborescente (no
reticular). A lo largo de los años y de las revisiones, Darwin introdujo otras
ideas externas (Lamarck, Geoffroy Saint-Hilaire, Von Baer) es su constructo
teórico, en buena parte como respuesta a las críticas recibidas desde el
colectivo científico de la época, y que en mi opinión añaden buena carga de
incoherencia a sus postulados iniciales sin resolver realmente dichas críticas:
herencia de caracteres adquiridos por acción directa (respuesta orgánica) e
indirecta (nicho formativo) de las condiciones del medio, teoría de la
pangénesis, etc. En síntesis, según el propio Darwin, el darwinismo clásico
podía resumirse en una serie de leyes que son: “la de
crecimiento con reproducción; la de herencia, que casi está comprendida en la
reproducción; la de variación por acción directa e indirecta de las condiciones
de vida y por el uso y desuso; una razón del aumento tan elevada, tan grande,
que conduce a una lucha por la vida y como consecuencia a la selección natural,
que determina la divergencia de caracteres y la extinción de las formas menos
perfeccionadas”.
Respecto al neodarwinismo, podemos considerarlo como una
purificación de las proposiciones del darwinismo clásico que rechaza la
pangénesis y la herencia de caracteres adquiridos, al tiempo que considera que
la fuente de las variaciones se encuentra en el “plasma germinal” (gametos) que
es ajeno a las modificaciones somáticas y a las señales que éste recibe del
medio. Su formulación se atribuye fundamentalmente a August Weissman, quién se
ha hecho célebre a partir de sus experimentos en los que mutilaba el rabo a ratas
de laboratorio para falsar la herencia de esta “modificación”. Aunque a día de
hoy pueda resultar sorprendente, no es de extrañar que estos experimentos no
tuviesen una gran aceptación en su época, dado que difícilmente puede
considerarse una mutilación como un carácter adquirido, y de hecho el
neolamarckismo se consolidó y continuó siendo una corriente importante durante
varias décadas posteriores. En cualquier caso, creo que en neodarwinismo puede
ser definido sin problemas como una teoría que reconoce a la selección natural
como el único “mecanismo” capaz de producir la evolución (gradual) de los seres
vivos.
Por último, la síntesis moderna (también llamada teoría sintética
o síntesis neodarwinista) puede considerarse como la unificación de la teoría
neodarwinista y la teoría mendeliana de la herencia bajo la forma de la
genética de poblaciones, iniciada por Fisher, Write, Haldane y Dobzhansky, y
que finalmente se extendería a otras ramas clásicas de la biología como la
paleontología (Simpson) o la zoología (Mayr, Huxley), y finalmente al conjunto
de las ciencias de la vida. La síntesis incorpora la genética como base de la
evolución, otorgando a las mutaciones espontáneas y “azarosas” el papel de
“fuente primaria de variación” y tomando las segregaciones mendelianas como
fundamentación de los procesos de la herencia. La selección natural es la
encargada (en sentido metafórico) de generar una reproducción diferencial
basada en la expresión de los caracteres codificados determinísticamente en los
genes y la supervivencia de los individuos con estas variantes (alelos) más
adecuadas. Como consecuencia se da un “cambio en las frecuencias alélicas de la
población” que favorece a los individuos con mayor “eficacia biológica”. Tal
como ocurría con el neodarwinismo, la síntesis rechaza cualquier influencia del
ambiente en la variación del material (genético) de partida, en frecuencia y
posición (asociado al neolamarckismo), así como el cambio saltativo
(mutacionismo) y las tendencias internas (ortogénesis). No obstante, a
diferencia de éste, la síntesis considera que existen otras procesos evolutivos
secundarios (de genética de poblaciones) que se escapan a la selección natural
y que entran dentro de la deriva genética.
Sin embargo, como cualquier neodarwinista moderno podrá ratificar,
existen numerosas excepciones, vacíos y ampliaciones al modelo original de la
síntesis. Tantas que algunos lo consideramos absolutamente insostenible a día
de hoy.
4-¿Puede resumirse la teoría darwinista actual de la evolución
como adaptacionismo más continuismo?
No. Este sería un resumen deficiente ya que sólo define parte de
la teoría. De hecho, el concepto de continuidad ni siquiera es patrimonio del
darwinismo, sino que es una de las tesis centrales del evolucionismo per se.
Con independencia del ritmo y el modo en el que se den las transformaciones, el
evolucionismo sostiene que la biodiversidad a través de los tiempos es producto
de una continuidad biológica que se opone a la aparición de novo de
formas sin ningún tipo de conexión filogenética (ya sea vertical u horizontal)
como explicación axiomática. Respecto al adaptacionismo, ciertamente esta es
una característica del darwinismo, ya que elimina explícitamente la posibilidad
de tendencias internas (ortogénesis y tendencia a la complejidad) y minimiza el
efecto de fenómenos no adaptativos (estructuralismo y neutralismo) en la
dirección de los cambios evolutivos, que responden fundamentalmente a un ajuste
de las características de los seres vivos al ambiente cambiante. Sin embargo el
adaptacionismo también es un componente clave del neolamarckismo clásico
(1860-1930), aunque como es fácil de entender los mecanismos propuestos son
diametralmente opuestos.
Si tuviese que resumir el pensamiento darwinista en una ecuación
simple, esta sería la siguiente: individualismo más competencia igual
reproducción diferencial. Por individualismo contemplo la singularidad única de
cada individuo y de sus variaciones, mientras que por competencia incluyo las
relaciones de exclusión entre individuos potencialmente viables por el acceso a
un recurso dado. Las dos condiciones son estrictamente necesarias para que se
de el único proceso de transformación poblacional que contempla el darwinismo.
Entiendo que es esta simple fórmula contiene la esencia de lo que se designa
como evolución por selección natural, y por ende, la esencia del darwinismo.
5-¿Cuál es la unidad básica de la evolución? Es decir, ¿qué es
exactamente lo que evoluciona: los genes, los organismos, las poblaciones, las
especies o todos ellos?
Bueno, aquí hay opiniones para todos los tipos. Algunos autores
tienen una respuesta fácil, rápida y aparentemente contundente al problema:
todas son unidades seleccionables y por tanto sujetos de la evolución.
Establecemos una teoría jerárquica de la evolución por selección natural (Gould
2004) y todos contentos.
En buena parte, esta diversidad es producto de unas definiciones
bastante laxas (por no decir incompletas o ilógicas) de todas estas categorías,
así como del paradigma neodarwinista que pone énfasis en la individualidad y la
selección competitiva. Creo que es necesario partir de una definición formal de
los sujetos y objetos a estudiar en Biología Evolutiva. En esto consiste una
parte importante de mi tesis, pasando por un cambio de enfoque hacia la
Biología Evolutiva de Sistemas.
6-También he encontrado en la literatura una sorprendente
flexibilidad en torno al concepto de "azar", tal como se usa en la
teoría sintética para las mutaciones genéticas. Algunos autores definen el azar
como " a-causal, sin causa alguna", otros como "lo que es
impredecible" (lo cuál puede representar simplemente una limitación
cognitiva del ser humano para predecir eventos que en el fondo sí tienen causas
reales), otros como "independientemente de que sea útil o no al
organismo". Todos estos conceptos de azar son diferentes e implican cosas
diferentes, es decir, no son equivalentes. En tu opinión, ¿qué significa
realmente, en la teoría sintética, que las mutaciones genéticas sean "al
azar"? ¿Hay alguna evidencia científica a favor de esa idea?
Lo que entiendo por azar en la síntesis moderna es el antagonismo
a la modificación del material hereditario inducida por el ambiente. Es la
negación de un papel activo de los individuos en su propia evolución y la
reciprocidad no-lineal entre organismo y medio. En esencia, es la exorcización
del lamarckismo, del evolucionismo primigenio. Y existen muy buenas razones
para que los neodarwinistas (a diferencia de Darwin) se opusieran frontalmente
a las tesis lamarkistas. El lamarckismo contempla un tipo de evolución no
selectiva, coordinada, que induce una transformación de las poblaciones a
través de los individuos que cambian al unísono. El proceso de homogeneización está implícito en el
mecanismo de cambio. La selección no solo es innecesaria, sino que es
imposible. En consecuencia, la variación azarosa e individual es una
imposición lógica del darwinismo.
Aunque la genética del siglo XX se construyó en gran parte en base
a unas premisas de cambio azaroso y una expresión unidireccional de la
información (Dogma central de la biología molecular), lo cierto es que
numerosas evidencias apuntan justo al lado contrario. No solo está más que
demostrado que los procesos de mutación suelen estar asociados a lesiones del
DNA, siendo los organismos los responsables de su aparición a través de los
mecanismos de reparación bioquímica, quedando patente que es el medio un agente
inductor de cambios a un nivel nucleotídico. Existen un buen número de
mecanismos de cambio molecular hereditario que están íntimamente relacionados
con las condiciones del medio y las respuestas que los organismos desarrollan
frente a estas. Estos cambios, que incluyen movilización de elementos móviles,
recuperación de las capacidades infectivas de virus endógenos, transferencia
horizontal de material genético y herencia epigenética de varias clases (marcas
en la cromatina, proteínas tipo prión, estructuras autorreplicativas, etc.), y
en todos ellos se puede observar un desequilibrio probabilístico de rango
variable (desde especificidad muy alta hasta espectros de variación más
relajados, pero en ningún caso puramente estocásticos) tanto en el su
acotamiento temporal como posicional (loci) en función a desencadenantes
ambientales específicos.
7-¿En qué consiste exactamente el concepto de "selección
natural" en la teoría sintética?
Como indiqué en la respuesta a la pregunta 3, la selección natural
es en la teoría sintética el “mecanismo” (por darle alguna categoría) capaz de
modificar la distribución de las frecuencias alélicas, entendiendo que dichos
alelos se han originado previamente por mutación al azar. Es por tanto el tamiz
que filtra las mutaciones y el timón que dirige la evolución de las especies, a
expensas del motor de las mutaciones, hacia los picos adaptativos. Es la mano
invisible del mercado del mundo natural.
8-Los neo-darwinistas mencionan como ejemplos recientes del poder
explicativo de la selección natural los siguientes: 1-El ejemplo del gusano de
la mosca de la manzana, que originalmente se alimentaba solamente de las frutas
del espino, pero que luego se adaptó a la manzana, y desde hace unos 100 años
comenzó a convertirse en una plaga importante en Estados Unidos. 2-La
resistencia por parte de cientos de especies de insectos a varios insecticidas
químicos, que hace muy fuertes a dichos insectos y muy difíciles de controlar.
3-La resistencia a los antibióticos. ¿Estos procesos son una prueba del poder
explicativo de la selección natural en los fenómenos biológicos y evolutivos?
En primer lugar, describir un fenómeno no es describir sus causas.
Estos ejemplos, que a mi me conste, no son una evidencia de la selección
natural sino del fenómeno de adaptación. Hoy sabemos que los mecanismos de
adaptación epigenética y los efectos transgeneracionales pueden tener un alto
potencial explicativo para algunos fenómenos adaptativos, especialmente los
relacionados con la alimentación. Esta es otra posible explicación a, lo que
insisto, es un la descripción de un fenómeno y no una prueba fehaciente de un
mecanismo o proceso.
En lo que respecta a la cada vez más distribuida resistencia a
antibióticos, parece que el principal mecanismo detrás de este fenómeno es la
transferencia horizontal. No podemos olvidarnos que las circunstancias altamente
nocivas o estresantes pueden detonar la potenciación de mecanismos
transferencia y de hipermutación. La capacidad de sobrevivir a altas dosis (y
resalto esta puntualización) de antibióticos gracias a la adquisición de
paquetes de información genética con disponibilidad poblacional difícilmente
puede ser invocado como prueba de la selección natural.
9-Muchos críticos del neo-darwinismo han argumentado que el
concepto de "selección" es en sí mismo inadecuado y engañoso, porque
presupone implícitamente un seleccionador (alguien que haga la selección o
escogencia entre las opciones disponibles) y por ende de un agente mental o
racional. Irónicamente, los darwinistas, son los primeros en negar la
existencia de una agencia mental en el proceso evolutivo, pero continuamente
emplean metafóras, terminos y ejemplos antropomórficos (que aluden a
características humanas) en la naturaleza para apoyar su posición, tales como
la idea de "El Relojero Ciego" o "El Gen Egoísta" y hasta
"La Madre Naturaleza", y el propio Darwin basó su idea de la
selección natural en la cría de animales domésticos, que es un ejemplo típico
de diseño y selección inteligente, es decir, que involucra una mente racional
que hace la selección o escogencia. En ninguna otra ciencia natural parece
ocurrir una paradoja similar. En tu opinión, ¿por qué los biólogos
neo-darwinianos, tan hostiles a cualquier idea de un diseñador inteligente en
la naturaleza, hacen uso continuo y reiterado precisamente de metáforas,
modelos, ejemplos y conceptos que presuponen implícitamente un diseñador o
seleccionador con características humanas?
No soy sociologo ni historiador, así que no me encuentro en
disposición de dar una respuesta realmente sólida. Supongo que existen varios
factores. Uno importante debe ser la marca de inicio del propio darwinismo. La
selección natural fue descrita como una metáfora. En realidad todo el “Origen
de las especies” está plagado de especulaciones y anécdotas muchas veces
inconexas. La visión de Darwin nunca fue sistemática (a diferencia de Lamarck),
y eso creo que ha marcado unos precedentes hasta hoy en lo que se refiere al
uso del lenguaje simbólico excesivo. En todo caso, también considero que el
concepto de selección natural es en sí mismo una trampa semántica, ya que el hecho
de seleccionar sugiere la necesidad de un seleccionador.
10-En las ciencias naturales las explicaciones son principalmente
de tipo causal y legal, es decir, se busca un mecanismo causal incluido una ley
natural que explique un fenómeno particular o conjunto de fenómenos. Así por
ejemplo la rotación terrestre explicaría causalmente el fenómeno de la
alternancia del día y la noche, y la rotación misma sería explicable por leyes
físicas. En el caso de la biología de la evolución, el fenómeno a explicar es
la evolución, y la selección natural se propuso originalmente como la
explicación de ese fenómeno (al menos así lo afirman muchos biólogos
actualmente). Sin embargo, para mi sorpresa, he escuchado a algunos darwinistas
actuales proponer que la selección natural no es la causa, sino el
"efecto" de la supervivencia diferencial de los organismos dentro de
las poblaciones (argumento que esgrime, por ejemplo, un darwinista en un debate
que sostuvo con Máximo Sandín). Ahora bien, si la selección natural no es la
causa sino el "efecto" de la supervivencia diferencial, entonces
¿cuál es la explicación científica que los darwinistas proponen como mecanismo
causal y legal de dicha supervivencia diferencial? ¿Y que quedaría entonces del
papel explicativo y casi omnipotente que juega la selección natural en la
teoría sintética?
El problema es que, en mi experiencia, ni los darwinistas tienen
demasiado claro qué es la selección natural. ¿Es un mecanismo? ¿Es un proceso?
¿Es un principio? ¿Es una consecuencia? Aunque la he visto citada como todo
esto, la selección natural no entra realmente en ninguna de estas categorías.
La selección natural es un término confuso que ni siquiera Darwin usaba de
forma concreta. Tal como resalta Wallace en una de sus cartas al mismo, éste
usa el término con dos sentidos distintos: “(1) para la simple preservación
de las variaciones favorables y la eliminación de las desfavorables, en cuyo
caso es equivalente a “la supervivencia del más apto”; y (2) para el efecto
producido por dicha preservación”. Por lo tanto, este es un problema de
base.
Creo que realmente la mayoría de los darwinistas nunca se han
planteado esta cuestión. Probablemente muchos se llevarían una buena sorpresa
al ver que sus cimientos son tan poco sólidos y absolutamente insostenibles
desde un punto de vista epistemológico como el que planteas.
11-Una objeción común al neo-darwinismo es que el registro fósil
no contiene los millones de formas transicionales que cabría esperar si la
teoría sintética fuese correcta. Los Darwinistas típicamente responden con, al
menos, tres argumentos: 1)El registro fósil no es perfecto; 2) Es muy difícil
encontrar los fósiles (el punto 1 y 2 explicaría la ausencia de muchas de esas
formas intermedias en el registro fósil conocido) y 3)Sí se han encontrado
formas intermedias o transicionales tales como el "Archaeopteryx",
que es una transición entre los reptiles y los pájaros modernos. ¿Qué opinas de
esta respuesta neo-darwiniana? ¿Es adecuada para responder a las críticas contra
el neo-darwinismo basadas en el registro fósil?
Las dos primeras son ciertas. Pero con importantes matices.
Evidentemente el registro fósil no contiene todas restos de todas las formas de
vida pretérita y es lógico que existan vacíos de conocimiento, dado que gran
parte del material se pierde por los procesos tafonómicos. Del mismo modo, los
fósiles no se encuentran distribuidos por doquier, es decir, existen ciertas
dificultades para encontrar material paleontológico. Pero el diablo vive en los
detalles, y he aquí como los matices son cruciales. En primer lugar, dado que
asumimos que el registro es imperfecto, la pregunta siguiente es si éste es
“representativo”, es decir, si aún con sus carencias refleja una parte
suficientemente importante de la evolución de la vida como para poder hacer
generalizaciones. Y los estudios de las últimas décadas indican que si lo es. Y
en segundo lugar, aunque los fósiles no se encuentran fácilmente en cualquier
lugar, existen lugares en los que se encuentran fácilmente. Los grandes
yacimientos paleontológicos (como Burguess Shale) ofrecen un registro bastante
detallado de los ecosistemas antiguos. Del mismo modo, también hay grupos cuyo
registro es considerablemente bueno, y es posible tener un reflejo bastante
nítido de la evolución de estos organismos. Curiosamente, ambos tipos de
registro favorecen la teoría del equilibrio puntuado, que se opone al
gradualismo filético propio del darwinismo al tiempo que ofrece una observación
que es fácilmente absorbible por fenómenos de evolución rápida y saltativa
concentrada en períodos cíclicos.
Respecto a las formas transicionales se pueden hacer varios
comentarios. Primero, la existencia de una continuidad es algo esperable de un
registro evolutivo con independencia del tiempo y los mecanismos que hayan
estado implicados en el mismo. Lo sorprendente sería no encontrar ningún tipo
de relación entre organismos, pues esto diría mucho en contra de la realidad de
la evolución como hecho. Lo que se pone en duda no es la existencia de conexiones
filéticas entre fósiles, sino los mecanismos y los procesos por los cuales se
ha originado esa relación. Dado que no podemos viajar en el tiempo, no podemos
inferir gran cosa sobre esto último. Lo que podemos cuestionar, y cuestiono, es
la gradualidad y la velocidad de estos cambios, dado que en caso de que los
cambios evolutivos tuviesen un recorrido primordialmente gradual el número de
fósiles transicionales excedería con creces al de las morfoespecies estables,
cosa que no ocurre. Y segundo, creo que buena parte de la interpretación de los
“eslabones perdidos” está sujeta a una preconcepción del resultado. Por
ejemplo, la evolución de los tetrápodos ha sido sistemáticamente descrita como
una serie gradual (pasando por Panderichtys, Tiktaalik, Acanthostega
e Ichtyostega) en la que se iban introduciendo los nuevos fósiles de
manera conveniente a la predicción gradual del modelo teórico. Sin embargo, muy
recientemente esta reconstrucción preconcebida ha resultado sólidamente
refutada debido a las incoherencias temporales entre estos fósiles, sus
reconstrucciones biomecánicas e icnitas de marcha cuadrúpeda dejadas por un
organismo indeterminado que preceden a todos los restos óseos de la serie
ideal. Esto indica una situación de solapamiento temporal entre todas estas
formas que en ningún caso obedece a una relación filogenética lineal y
gradualmente ordenada.
12-Para formular y entender correctamente la siguiente pregunta,
me gustaría que los lectores de esta entrevista analicen los gráficos
siguientes sobre el reino animal y algunos de sus filos y subfilos:

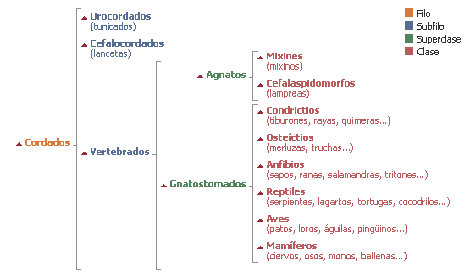
Basado en la anterior clasificación, algunos críticos del
neo-darwinismo sostienen que, aún si fuesen aceptables como evidencia
científica los típicos ejemplos de evolución y las formas transicionales que
proponen los darwinistas (por ejemplo, las transiciones entre los reptiles y
los pájaros, la evolución del caballo, etc.) tales ejemplos constituirían meras
trivialidades en comparación con lo que la teoría sintética tendría explicar
mediante la selección natural y las mutaciones al azar. Tales ejemplos (la
evolución del caballo, por tomar un caso), típicamente, son casos de evolución
dentro de un mismo filo o subfilo (ej: los vertebrados), que es prácticamente
insignificante en comparación con el árbol completo de la vida. La pregunta es:
¿No está la teoría evolutiva actual basada en una inmensa e injustificada
extrapolación de una parte mínima de la realidad biológica a todo el inmenso
conjunto de la vida? ¿Por qué tienen tanta confianza los darwinistas en el
poder explicativo de la selección natural, cuando la evidencia científica
disponible actualmente sólo explica, como máximo, una muy minúscula parte del
inmenso arbol de la vida? ¿Qué evidencia científica sólida hay, por ejempo, de
que un murciélago y una esponja de mar hayan evolucionado por selección natural
y mutaciones al azar a partir de un ancestro común?
Realmente lo primero que habría que comentar es que la iconografía
del árbol de la vida no es sostenible por más tiempo, al menos según nos
indican los datos moleculares. En realidad, las nuevas filogenias contienen un
profuso número líneas horizontales, que responden a fenómenos de hibridación,
reticulación introgresiva, transferencia horizontal y adquisición de virus y
elementos móviles. Estas relaciones filogenéticas de carácter no vertical se
dan a lo largo de todos los reinos, desde las bacterias a los primates. De
hecho, en el primer caso, el volumen de material genético que ha sido sujeto de
transferencia horizontal en algún momento supone más de un 80% de media de los
genomas procariotas. En el segundo ejemplo, por solo citar un par de ellos, se
han encontrado evidencia de hibridación entre géneros de primates, dando lugar
a especies híbridas, así como por supuesto la entrada puntual de nuevos
elementos móviles por transferencia horizontal e inserciones retrovirales en
los genomas del grupo. Y esto no es ninguna excepción. En algunos grupos, como
las angiospermas, la reticulación y la hibridación son por el contrario la
norma. Esto ha llevado a redibujar el esquema de la vida, no de una forma
arborescente como la de las heráldicas que recoge el darwinismo, sino como un
tapiz entretejido con unas líneas maestras (las relaciones de herencia
vertical) y infinidad de hilos que lo cruzan (las relaciones de herencia
horizontal).
A partir de este punto es fácil ver que las filogenias más
modernas no se ajustan a las predicciones del darwinismo. Si a esto
incorporamos la naturaleza puntuada y los saltos del registro fósil el panorama
no puede ser más distinto. Para mas inri, los ejemplo de transición gradual no
son todo lo claros que debieran. Ya he expuesto el problema en la respuesta a
la pregunta anterior, pero podemos dirigirla al ejemplo concreto de la
“evolución del caballo”. La secuencia de fósiles en esta filogenia no es
realmente lineal, sino que se trata de una ordenación ideal de estadios entre
restos que no mantienen una relación temporal equivalente. Es una
reconstrucción ideal. Por lo tanto, no es sólo que el darwinismo esté extrapolando
un ejemplo minoritario a la totalidad del registro fósil, sino que ese ejemplo
es una construcción teórica. Las razones de esa confianza absoluta en el poder
de la selección me son ajenas, aunque sospecho que tienen que ver con el
“adoctrinamiento” en el proceso formativo de los biólogos, la inercia de los
paradigmas científicos y el refuerzo (positivo y negativo) que recibimos desde
el resto del colectivo científico, laboral y socioeconómico.
En todo caso, del mismo modo que exponía en la respuesta anterior,
no se pueden tomar los restos fósiles como prueba de los mecanismos evolutivos,
tan solo de los fenómenos. A partir de ahí se debe encontrar una coherencia
entre la teoría y las pruebas históricas, pero esta relación no es
bidireccional. No hay ninguna prueba de que las esponjas y los murciélagos
hayan evolucionado por selección natural, porque no existe un registro
demográfico del proceso. Lo más que podemos hacer es complementar las pruebas
paleontológicas con datos neontológicos (moleculares y anatómicos) para tratar
de obtener pistas sobre los mecanismos moleculares de cambio implicados en el
fenómeno. De nuevo, aquí tampoco encontramos una prueba de la selección natural
en sí. Lo que podemos observar es que la mutación puntual sobre los “genes”,
las secuencias codificantes de proteínas que se encuentran en el motor de la
síntesis moderna, no pueden explicar la evolución de estos organismos debido a
la extensa homología y la falta de innovación en estas secuencias que se
encuentra a lo largo de las comparaciones filogenéticas desde el hombre hasta
las esponjas. Por el contrario existen
claras evidencias de la importancia otros mecanismos como la reordenación de
los cromosomas, la duplicación de genomas y secuencias genéticas, la
adquisición de material genético foráneo y la reorganización física y funcional
de la información por medio de elementos móviles.
13-Otra objeción contra el neo-darwinismo es que la idea original
de "la supervivencia del más apto" es tautológica, es decir, es una
afirmación verdadera en virtud solamente de su forma lógica pero vacía de
contentido empírico y predictivo, porque se define a los más aptos como
aquellos que "sobreviven" (o dejan más descendencia) y a los que
sobreviven (o dejan más descendencia) como aquellos que son más aptos. Muchos
darwinistas se han dado cuenta de este problema, y han reformulado la idea
diciendo que la seleccion natural es realmente una diferencia en el éxito
reproductivo, concepto que incluye tanto la habilidad de sobrevivir hasta la
edad reproductiva como la capacidad de reproducirse. ¿Qué opinión te merece
esta reformulación darwinista?
Bueno, es una salida por la tangente, pero en el fondo no deja de
ser una definición tautológica maquillada y, como ya he comentado, mal
planteada. Dado que este es un problema de base (la inexactitud categórica y
terminológica del darwinismo) hemos llegado a una situación en la que podemos
jugar a nuestro antojo con las definiciones, hasta el punto de darle a la
selección natural el carácter que nos parezca más oportuno según las
necesidades a cubrir.
Por otra parte, esta formulación sigue sin satisfacer ni siquiera
las bases de lo que entiendo por pensamiento darwinista, dado que “una
diferencia en el éxito reproductivo” puede darse por factores ajenos a la
selección competitiva, a través de sesgos de viabilidad (restricciones
facultativas) e incluso por pura contingencia.
En cualquier caso, creo que el término selección natural es un
pesado lastre para la biología y tratar de definirlo es tan innecesario como
improductivo, tanto por su inexactitud como por su falta de contenido.
14-También se ha criticado al neo-darwinismo que, como teoría, no
tiene capacidad predictiva. Pero los darwinistas responden que sí tiene
capacidad predictiva, y que eso se resume en la siguiente predicción: en un
contexto ambiental específico, un genotipo va a ser más exitoso que otro en su
supervivencia o en su reproducción por razones que tienen que ver con el medio
ambiente o con los otros organismos dentro de la población. ¿Qué opinas de
esto?
Ciertamente, no tiene capacidad predictiva. Esto ha sido admitido
al menos por algunos biólogos darwinistas, como Conways Morris, y no deja de
ser una limitación importante para una teoría científica. Por supuesto, gran
parte del material de estudio de la evolución es de carácter histórico
(fósiles, filogenias moleculares, anatomía comparada, etc.) lo que llevan a un
análisis a posteriori y a la idea de que “la biología evolutiva es una
ciencia histórica”. El darwinismo tan sólo ofrece predicciones muy vagas (por
llamarlas de algún modo) al adjudicar el origen de las analogías a las
presiones selectivas del ambiente.
No obstante, mi impresión personal es que la evolución obedece a
mecanismos, procesos y dinámicas
científicamente describibles, y estas deben de ser descritas y estudiadas por
la Biología Evolutiva. A partir de aquí podríamos hablar de fenómenos de cambio
probabilísticamente predecibles. Y digo probabilísticamente porque al hablar de
sistemas complejos (como son los sistemas biológicos) entramos frecuentemente
en el dominio del caos. Sin embargo, aún en sistemas de este tipo es posible
obtener resultados de carácter probabilístico con cierta capacidad de
predicción, algo que supera con creces las aportaciones actuales de la
síntesis.
Respecto a la supuesta capacidad predictiva que enuncias, bueno,
no me parece que esto pueda llamarse como tal. Es una explicación a
posteriori, y una tesis no falsable ni demostrable, no una predicción. De
hecho, me gusta exponer este problema con un ejemplo culinario. Imaginemos que
se tiene un conjunto de peras y manzanas en un momento inicial t=0, con iguales
proporciones. Ahora saltemos hasta un momento temporal posterior, t=1, y
recontemos la proporción de cada fruta. Lo cierto es que no importa si sólo
quedan manzanas o peras, en cualquier caso podemos afirmar que han sobrevivido
los más adecuados frente a un agente selectivo (en este caso un humano
hambriento). Pero lo más interesante es que, aunque queden manzanas y peras en
proporciones variables, siempre podemos afirmas que las que quedan son las más
adecuadas de entre su tipo. Lo que quiero ilustrar con este ejemplo tan básico
es que, a posteriori, es fácil ajustar los resultados a “las
predicciones de la teoría”, cuando la única predicción es la supervivencia de
los mejor adaptados. La supervivencia no es el cambio y por lo tanto no es una
explicación de base a los fenómenos evolutivos.
15-En su libro "What Darwin Got Wrong", el filósofo y
científico cognitivo Jerry Fodor y el biólogo molecular Massimo
Piattelli-Palmarini (ambos materialistas, ateos y anti-creacionistas) afirman
que la teoría sintética de la evolución no es una teoría científica, sino una
forma de narrativa histórica. A diferencia de las teorías científicas, que
predicen un fenómeno concreto basado en leyes naturales, la teoría sintética de
la evolución es más bien una reconstrucción, a posteriori, de cómo los
organismos se pudieron haber adaptado mediante la selección natural. ¿Compartes
esta opinión?.
Absolutamente. Creo que esto resume a la perfección lo que quería
transmitir en la respuesta a la pregunta anterior.
16-¿En qué consiste el nuevo modelo teórico que biólogos
innovadores como tú proponen y qué ventajas tiene en relación con la teoría
sintética?
Actualmente no son pocos los biólogos que encuentran una
importante insatisfacción con la síntesis moderna y buena parte del aparato
teórico que implica (determinismo genético, linealidad, totipotencia de la
selección natural, gradualismo, mendelismo, etc.) Esto nos sitúa en un momento
tan interesante como caótico (en el sentido tradicional) donde existen varias
proposiciones inconexas que vienen a remendar las carencias de la síntesis a
través de ampliaciones y de la inclusión selectiva de excepciones. Estos
modelos amplían el estrecho cerco de la síntesis moderna por medio de la
epigenética, la genómica, la biocomputación, la microbiología y la embriología,
por citar algunos campos, pero sin modificar el núcleo teórico del darwinismo.
Sin embargo, estas visiones no están integradas entre sí y arrastran todos y
cada uno de los lastres conceptuales del darwinismo que he ido exponiendo a lo
largo de la entrevista. Esto en mi opinión, y en la de otros científicos con
mayor recorrido que el mío, esto es un error. No vale con remendar el
darwinismo, hay que reformular la Biología Evolutiva desde un enfoque teórico
diferente que sea capaz de dar una explicación coherente a los principales
problemas de la síntesis sin perder poder explicativo para el resto de fenómenos
descritos.
Siguiendo esta línea, a mi juicio la alternativa más potente
planteada hasta la fecha, tanto por su capacidad explicativa como por su
monumental cambio de concepción hacia la biología, es la “Teoría de la
evolución por integración de sistemas complejos” de Máximo Sandín.
Máximo
Sandín rechaza y critica abiertamente al darwinismo en su aspecto científico y
en su nefasta influencia social, propone una visión alternativa de la vida y de
la evolución que, en contraste con competencia radical de la síntesis, se basa
en la integración de sistemas vivos. La adquisición, la modificación y la
compartición de la información contenida en, y a través de, bacterias y virus –
sistemas biológicos más básicos – por parte de otros organismos y entre sí, son
los principales procesos que originan la novedad evolutiva. La transferencia
horizontal, la endosimbiosis y, muy especialmente, las dinámicas de los virus y
secuencias afines son, para Sandín, los
últimos responsables de los cambios de organización que se observan en el
registro fósil, de carácter puntuado, que se producen con cierta periodicidad y
a continuación de las extinciones en masa. Estas extinciones han sido asociadas
por varios autores con momentos de profunda crisis medioambiental, y Sandín considera
que estas crisis no solo constituyen un sesgo sobre la biodiversidad, sino
también un detonante para la innovación. La desestabilización profunda del
medio puede inducir la movilidad de los elementos móviles del genoma y de los
virus endógenos, así como potenciar la
transferencia horizontal, y remodelar los ecosistemas de manera rápida a través
de cambios en las especies de carácter saltacionista. Estos cambios, en
consecuencia a las dinámicas de estos mecanismos, no son azarosos, sino
fuertemente limitados y dirigidos, y son responsables de la aparición paralela
de caracteres homólogos entre individuos de la misma especie, e incluso, entre
especies distintas. El cambio evolutivo, por lo tanto, es presentado por Sandín
como un fenómeno no competitivo sino integrador, que está modulado por el
ambiente y produce respuestas acotadas, que acontece sobre el conjunto
poblacional en vez de sobre el individuo aislado, y que no es consecuente con
un patrón gradual y de ritmo constante, sino puntuado y con tendencia a crecer
en complejidad. Además, la adaptación, la especiación y la generación de
verdaderas novedades o apomorfías (que él denomina transformación), son
aspectos desacoplados (aunque relacionados) de la modificación de las especies
en el tiempo.
Mi propuesta personal consiste en una reformulación integral de la
Biología Evolutiva a través de la integración de las ciencias de la complejidad
con las ciencias biológicas bajo el paradigma sistémico para generar un nuevo
catálogo de categorías, herramientas conceptuales y dinámicas concretas que
amplían, potencian y dan consistencia al estudio de la evolución a través de un
nuevo área de conocimiento que identifico como Biología Evolutiva de Sistemas.
Por supuesto, esta propuesta toma como eje teórico el modelo de Máximo Sandín,
al tiempo que explora nuevas fronteras, formaliza los términos y trata de
unificar de manera no sólo coherente, sino inseparable, a buena parte de los
fundamentos heterodoxos clásicos (predarwinistas y presintéticos) y de los nuevos
retos de la biología del siglo XXI, en una alternativa potente a la síntesis
moderna y al pensamiento darwinista. Por otro lado, recalco el uso de nuevas
herramientas de biocomputación para el estudio de la Biología Evolutiva de
Sistemas, y actualmente estoy trabajando en un modelo simulado que trata de
reproducir mis propuestas más concretas en un escenario virtual. Y hasta aquí
puedo leer.
17-En tu nuevo modelo teórico, ¿cuál sería el rol explicativo de
la selección natural, de la deriva genética y que papel jugaría la genética de
poblaciones?
Entiendo por selección natural aquellos fenómenos de supervivencia
diferencial mediados por la competencia entre organismos por lo demás viables.
Esta es una puntualización que sin duda no gustará a muchos darwinistas, dado
que acota el poder adjudicado a este término, pero considero que esta es la
innovación del concepto tal como lo concibió Darwin y sus precursores, dado
que la idea de una supervivencia acotada
a lo largo de la historia de la biodiversidad precede con mucho a Darwin, y la
verdadera innovación consiste en la idea de que la competencia es lo que lleva
a la selección. Esto excluye a todos aquellos fenómenos de restricción que
juegan con la viabilidad en términos absolutos de los organismos, ya sean de
corte ecológico (coherencia ecositémica) o embriológico (constricciones del
desarrollo). Para que se pudiera dar un fenómeno de este tipo, sería necesario
contar con un escenario en el que existiesen unos umbrales de competencia
suficientemente altos como para llevar a la muerte o la no reproducción de
algunos individuos en función de sus ligeras variaciones individuales. Existen
varios estudios sobre redes ecológicas que reflejan justamente lo contrario,
que los ecosistemas se autoorganizan evitando la competencia y la exclusión
competitiva. No obstante, para que esta selección competitiva tuviera una
continuidad temporal que lleve a la composición de las poblaciones, además de
un elevado umbral de competencia, los individuos implicados deberían de sobrevivir
en función de sus características y estas deberían de ser heredables fielmente
con el fin de que no se altere la supuesta ventaja que otorgan (fidelidad de
copia), dado que en este último supuesto no podría repetirse la selección a lo
largo de suficientes generaciones como para llevar a una homogeneización por
remplazo. Si consideramos los caracteres como elementos discretos, con
expresión lineal y determinista, y sin posibilidad de influencia recíproca
entre sí y con el medio, podemos mantener esta suposición como válida. Esto es
lo que supone la síntesis moderna al fusionar el neodarwinismo con el
mendelismo. Sin embargo, esta suposición no es sostenible a la luz de la
genómica actual. Solo un bajo número de caracteres presentan segregaciones mendelianas
(si no recuerdo mal, menos del 8% de las enfermedades humanas hereditarias,
casos muy estudiados, entrarían en esta categoría), mientras que la expresión
general de la información genética se realiza a través de redes complejas de
interacción, en las cuales es fundamental el contexto genético y ambiental como
conjunto. Esto sin contar con otros canales de información epigenética y
ambiental, que de ningún modo pueden atribuirse a las segregaciones
mendelianas, y a aquellos mecanismos de robustez que tienden a limitar las
variaciones en torno a estados multifactoriales estables y bien definidos.
Por lo tanto, entiendo que el concepto de selección natural haría
referencia a un fenómeno de reproducción y supervivencia diferencia mediada por
competencia, siendo un caso muy particular de las restricciones al cambio
evolutivo. Solo cuando los organismos reproductores deben su “éxito” a
caracteres heredables e individuales de forma determinista y las condiciones de
elevada competencia se mantienen durante suficiente tiempo, la selección
competitiva podría (y remarco el condicional) tener un efecto homogeneizante en
la población. En esencia, el papel de la selección natural, aunque teóricamente
posible, tiende a cero en un escenario natural (ajeno a las predisposiciones de
la experimentación en laboratorio).
Respecto a la deriva genética, este es un término que viene a
incluir a todos aquellos fenómenos de cambio en la composición de las
frecuencias alélicas de una población no producidas por selección, sino por
sesgos de población indiscriminados. Es una definición por oposición y, por lo
tanto, prefiero obviarla en mi análisis porque no aporta nada.
Ambos términos están en estrecha relación con la genética de
poblaciones. Personalmente rechazo esta disciplina como base de la Biología
Evolutiva y creo que aporta muchas más trabas que ayudas al estudio de la
evolución. En un momento como el actual, en el cual trabajamos con redes
genéticas y se nos escapa cuál es la unidad material de la información biológica,
es bastante improductivo estudiar la distribución de polimorfismos genéticos y
cómo estos varían con el tiempo. En vistas a lo anterior, no podemos establecer
una causalidad directa entre el genotipo y el fenotipo, y, aún en los casos en
los que es posible obtener cierta información, no salimos de una cuestión
esencialmente demográfica, microevolutiva en el mejor de los casos, que nada
tiene que ver con los cambios macroevolutivos patentes en el registro fósil y
la genómica comparativa.
18-En los medios de comunicación se lee o escucha muchas veces que
se ha descubierto el "gen de la homosexualidad", o el "gen del
alcoholismo" el "gen de la creatividad", o el "gen de la
infidelidad". Esto se basa en la idea del gen como una unidad discreta de
información genética responsable de una característica particular, pero he
leído que esta comprensión simplista sobre los genes está siendo puesta en duda
los datos científicos más recientes. ¿Puedes comentarnos algo al respecto?.
Sí, así es. Las “ciencias de la información biológica” (genética,
epigenética, genómica, etc.) están demoliendo literalmente las ideas clásicas
de la herencia y la expresión de la información. Pese a ello, no son pocos los
medios, e incluso las publicaciones científicas, que siguen haciendo uso de
conceptos laxos o incoherentes, en especial en lo que se refiere a enfermedades
y comportamientos humanos.
Por un lado, la información genética tiene una expresión y una
estructura harto compleja que no se ajusta a las concepciones lineales y
deterministas del siglo pasado. La composición de los genomas dista de ser una
ristra de “genes” (secuencias delimitadas de DNA, que se expresan
funcionalmente como proteínas y algunos RNA largos, y que actúan como unidades
discretas de información), ordenadas una detrás de otra a lo largo de los
cromosomas. De hecho las secuencias codificantes de proteínas, los
tradicionalmente llamados “genes”, constituyen una fracción minoritarias de los
genomas eucariotas (cerca del 2% del total en humanos, por ejemplo), al tiempo
que existe una enorme homología y conservación entre estas secuencias a través
de la filogenia que implican un papel secundario en la evolución de los
genomas. Del mismo modo, el número de “genes” tampoco es indicativo de la
complejidad final de los organismos (los seres humanos contamos con unas 25.000
de estas secuencias, mientras que la mosca de la fruta cuenta con unos 18.000 y
el arroz más de 30.000). Esto nos lleva a pensar que existe mucha más
información que la adjudicada a los “genes”. Además, estas secuencias distan de
tener una expresión determinista y lineal, dado que la mayoría están sujetas a
fenómenos de modificación postraduccional y postranscripcional, regulación por
promotores alternativos, marcas de splicing y de poliadenilación, enhancers,
etc., que dependen de las circunstancias del contexto celular y biológico. Del
mismo modo, las secuencias codifiantes tampoco tienen unos límites físicos bien
definidos, dado que los fenómenos de trans-splicing, quimerismo y solapamiento pueden
empalmar facultativamente secuencias no necesariamente colindantes. Por último,
es cada vez mas patente que existe una enorme diversidad de elementos sin
codificación a proteínas que se expresan en forma de multitud de tipos de
pequeños RNA con funciones reguladoras que se escapan a la definición de “gen”
y que se encuentran incluidas dentro de las complejas redes de interacción
genética que, insisto, son dependientes de los contextos genéticos y
ambientales. Como consecuencia, no es posible establecer una relación lineal
entre el “gen” y el carácter, mucho menos cuando estos son tan complejos como
un comportamiento humano. Por supuesto la mayoría de los genetistas no comunica
dicha relación actualmente (a diferencia de los medios), sino que hablan de una
“predisposición genética”. Sin embargo, tal como decía un buen colega y tutor,
en ausencia de un marco de referencia completo, se podría decir que los hombres
tenemos predisposición genética a ser mujeres, pues tenemos un cromosoma X. Lo
que intento decir es que la “predisposición” genética es una falacia en
ausencia de un conocimiento completo de todas y cada una de las variables que
se dan en un momento dado de la vida de un organismo.
Por otra parte, no podemos olvidarnos de que existen otros canales
de información que interactúan con el nivel genético, los canales epigenético y
ambiental. Por lo tanto, hablar de “genes de” me parece insostenible y
contraproducente. Y en todo caso, visto lo visto, ¿qué es un gen, si es que es
algo?
Este es un tema que sobre el que he escrito con algo más de
extensión en mi artículo “El mito del gen: genética, epigenética y el bucle
organismo-ambiente”, que incluyo en la bibliografía recomendada para aquellas
personas que les interese ampliar sobre el mismo.
19-Ante las propuestas innovadoras de biológos como tú o Máximo
Sandín, algunos neo-darwinistas afirman
que tales propuestas no refutan el papel esencial de la selección natural en la
evolución, sino que únicamente amplían y enriquecen la fuentes posiles de variación
genética. Por ejemplo, afirman ellos, un organismo o conjunto de organismos
originados por una duplicación, o por una reorganización genómica, o por una
transmisión horizontal de genes, o por simbiosis, deben en última instancia
enfrentarse a la prueba decisiva de la selección natural o
"filtración" por parte del ambiente, que es lo que en definitiva va a
determinar cuáles son los organismos que tendrán diferencialmente más éxito
reproducitvo y por ende predominarán en una población, por lo que la selección
natural seguiría siendo esencial en la explicación científica de la evolución.
¿Qué opinas de esta objeción?
Bueno, creo que he ido contestando a esta pregunta a lo largo de
la entrevista. En vistas a los modelos teóricos que trabajamos, la selección
natural no solo está mal definida y como término no aporta nada al estudio de
la evolución, sino que es innecesaria (y en gran medida insostenible) en
función a los mecanismos, procesos y dinámicas que introducimos.
20-Los críticos del darwinismo han analizado las posibles causas
de la hegemonía de esa cosmovisión a la luz de factores socio-económicos y
culturales. Ciertos críticos afirman que el darwinismo es, histórica e
ideológicamente, un apéndice de la teoría económica del libre mercado, que tiende
a apuntalar conceptos como los de "competencia" o "escasez de
recursos", que son conceptos fundamentalmente económicos-empresariales, y
que refuerzan las diferencias sociales dentro de una sociedad basada en ese
modelo económico capitalista, justificando así a las clases poderosas o
dominantes las cuáles serían, en la interpretación darwinista, las "más
aptas". ¿Estás de acuerdo con esta opinión?
Si, estoy de acuerdo. Aunque entiendo que la relación es
retrolimentada. El darwinismo se nutrió de las ideas de Malthus, Spencer, Adam
Smith y Hobbes. Es una cadena de pensamiento, una cosmovisión común que, creo,
queda sintetizada y convertida en ley natural a través del darwinismo. Esta
cosmovisión no solo incluye el libre mercado, sino que engloba en general una
idea de supremacía por derecho que permite el progreso de unos sujetos
privilegiados a través de la degradación y la explotación del resto, en lo que
perciben como una lucha natural de todos contra todos. Como decía, el
darwinismo se fundó sobre esas ideas (tal como relata el propio Darwin y muchos
historiadores modernos), aunque no cabe duda que inmediatamente el darwinismo
se convirtió a su vez en una herramienta, una justificación científica, que
apoyaba el nuevo orden de poder emergente que era el liberalismo económico de
la época victoriana. En consecuencia, “El origen de las especies” tuvo una gran
aceptación social (que no de los estamentos eclesiásticos ni del colectivo
científico) agotándose rápidamente la primera edición. Pero además, el darwinismo
fue promocionado a través del X-Club, la “más poderosa sociedad científica” de
la época, cuyos integrantes eran altos cargos científicos partidarios del
liberalismo, hasta el punto de fundar la revista “Nature” como púlpito del
darwinismo.
Es bien conocido que desde su popularización, el darwinismo ha
sido empleado como punta de lanza para llevar a cabo prácticas económicas de
explotación laboral, y que grandes magnates como Rockefeller no tenían pudor en
usar el darwinismo como una explicación natural a su privilegiada situación de
poder y riqueza. Del mismo modo, el darwinismo permitió dar un enfoque
científico, y por tanto justificable, a los prejuicios sociales y raciales. Es
más, estableció un nuevo escenario temporal en el cual se volvía una “obligación
moral” mejorar la especie humana mediante la dirección de la reproducción y la
supervivencia de los grupos humanos. Esto es lo que Francis Galton, primo de
Darwin, denominó como eugenesia. En mi opinión, la eugenesia ha sido el
movimiento sociopolítico más turbio e influyente en la historia de la biología,
y es imposible no ligar su desarrollo (una vez más retroalimentado) al
darwinismo. La puesta en marcha de programas eugénicos para la reproducción
concertada de los individuos “más virtuosos”, así como el aislamiento, la
esterilización y el asesinato de los “lastres sociales” (a los ojos de los
eugenistas, claro está) tuvo una repercusión en buena parte de los países
occidentales hasta el final de la segunda guerra mundial, aunque sin duda la Alemania
nazi lo llevo hasta el grado más alto en lo que se refiere a relevancia para el
estado. La eugenesia fue el motor ideológico y económico detrás de buena parte
del desarrollo de la genética clásica y de la síntesis moderna, y su expansión
ha configurado, en mi opinión, toda la biología del siglo XX y los mitos
sociales que han permeado hasta la sociedad, pasando por la defensa del
mercado.
21-Otros críticos argumentan que, si bien los factores
socio-económicos mencionados en la pregunta anterior explican la gran
aceptación y receptividad que la obra de Darwin tuvo al comienzo, en la
actualidad la hegemonía del darwinismo no puede explicarse en esa forma, entre
otras cosas porque ya nadie emplea el darwinismo para justificar las
diferencias entre las clases sociales, y porque la mayoría de biólogos
darwinistas son progresistas, simpatizantes del socialismo y críticos del libre
mercado. La hegemonía actual del darwinismo se explicaría mejor en términos de
la lucha cultural occidental entre la religión y el secularismo. Según estos
críticos, el darwinismo muestra que Dios no existe o cuya existencia es
innecesaria y superflua. Y dado que esto es la voz de la "ciencia",
entonces se tiene a mano el mejor argumento científico a favor del ateísmo y de
una cosmovisión secular. Esta hipótesis, dicen estos críticos, tiene un gran
poder para explicar lo siguiente: la tendencia de muchos darwinistas de
exagerar tremendamente los poderes creativos de la selección natural elevándola
casi al nivel de Dios (la selección natural sería el sustituto científico,
natural y ateo del Dios creador del universo, cumpliendo así el rol de un mito
secular); la obsesión de los darwinistas por interpretar cualquier crítica a la
selección natural como una defensa del "creacionismo" (que es la
reacción simplista propia de los miembros de una secta religiosa que se ponen
inseguros y a la defensiva cuando su doctrina criticada), o la tendencia casi
paranoide de ver a los críticos como "creacionistas encubiertos".
También explicaría la distorsión histórica según la cuál fue Darwin quien
ofreció por primera vez una explicación de la evolución en términos
naturalistas (cuando en realidad la evolución se venía estudiando y tratando de
explicar en forma naturalista desde hace casi 100 años antes). Y explicaría
también la famosa y influyente frase de Richard Dawkins: "Aunque el
ateísmo pudo ser sostenible en términos lógicos antes de Darwin, Darwin hizo
posible ser un ateo realizado intelectualmente" (esta frase de Dawkins
implica que, si la teoría de Darwin es falsa, la presunta sotenibilidad
intelectual del ateísmo se derrumbaría, algo que comprensiblemente y como
ocurre con cualquier creyente cuando le atacan su cosmovisión, los ateos
estarán dispuestos a evitar a toda costa, lo que explicaría la
hipersensibilidad que tienen a la crítica del darwinismo). ¿Qué opinión te
merece esta segunda explicación alternativa de la hegemonía darwiniana?
Me parece una explicación forzada. En realidad el debate entre
evolucionismo y creacionismo es una aberración histórica. Como otros han
señalado anteriormente, la teología y la ciencia son dos dominios de
pensamiento humano independientes. O al menos deberían de serlo. El punto inicial de este enfrentamiento puede
situarse en la polémica que desató el “Origen de las especies” en la jerarquía
eclesiástica de Inglaterra. Hay que señalar que en aquella época y lugar no
existía una separación clara entre ambos estamentos, y era frecuente que los
cargos universitarios fueran al tiempo cargos eclesiásticos. Por supuesto, esta
es una situación particular que no es equiparable con la actual. Entonces, ¿por
qué continua este enfrentamiento?
Mi impresión es que los darwinistas hicieron acopio de una de las
tácticas políticas más usadas de todos los tiempos: basar tu campaña en las
deficiencias del adversario. Y con el tiempo, esta práctica se ha vuelto tan
habitual que forma parte del “credo” darwinista hasta llevarlo a una situación
dogmática. Por otra parte, resulta curioso como se ha ido incorporando este
debate en un país como el nuestro, de larga tradición católica, en el que el
evolucionismo y la religión parecen haber convivido sin demasiados problemas
durante bastante tiempo.
Tachar a los críticos del darwinismo de “creacionistas
encubiertos” es, a mi juicio, un mecanismo de defensa que inmediatamente
convierte lo que no se entiende o no se comparte en una categoría tratable,
reconocible y sobretodo, atacable. Desde el otro lado, los llamados
“creacionistas” tratan de reforzar su fe entrando incluso en dominios que no
siempre controlan, al tiempo que tratan de utilizar cualquier vacío (y hay
muchos donde elegir) como una supuesta prueba de la necesidad de un poder
superior. Aunque pueda parecer una relación equilibrada, lo cierto es que la fe
es una decisión existencial y personal, y puede buscar soportes dónde sea dado
que no obedece a normas estrictas del mismo modo que no requiere de pruebas
fehacientes. Sin embargo, la ciencia debe de obedecer a una serie de normas
epistemológicas que deberían de evitar que se cayera en discusiones, a mi
parecer, tan absurdas como estas. En su casa que cada uno crea lo que quiera
(si es que quiere creer en algo) pero
el ejercicio científico debería de centrarse únicamente en el conocimiento
basado en la experiencia y en los hechos falsables.
22-¿Qué bibliografía y páginas web recomendarías a los estudiantes
de biología y a lectores en general para que profundicen más en los temas
discutidos en esta entrevista?
Recursos en la web:
- BiVOS. Grupo de Estudios en Teóricos en
Biología Evolutiva de Sistemas: http://bivos.cooperativakatal.org/
- Somos bacterias y virus. Página
personal de Máximo Sandín: http://somosbacteriasyvirus.com/
- Biología y pensamiento. Blog CSIC de
Emilio Cervantes: http://www.madrimasd.org/blogs/biologia_pensamiento/
- Evolución y ambiente. Página del
Instituto de Estudios de Evolución Humana: http://www.iieh.com/
En texto:
- Pensando la evolución, pensando la
vida. Máximo Sandín 2006. Cauac.
- La crisis latente del darwininismo.
Mauricio Abdalla, 2007. Cauac.
- La tautología darwinista. Fernando
Vallejo, 2002. Taurus.
- Darwinism: the refutation of a Myth.
Søren Løvtrup, 1987. Croom Helm.
-
Ecological Developmental Biology: Integrating Epigenetics, Medicine, and
Evolution. Scott F. Gilbert y David Epel, 2009. Sinauer.
- Evolución: la increíble historia de una
teoría científica. Edward Larson, 2006. Debate.
- El mito del gen: genética, epigenética
y el bucle organismo-ambiente. Revista de medicina naturista. Vol. 6, Nº 1, 2012 ,
págs. 42-49. http://dialnet.unirioja.es/servlet/articulo?codigo=3829210
- Modelos evolutivos predarwinistas.
Andrés Galera, 2002. Arbor. Vol 172, No 677. http://arbor.revistas.csic.es/index.php/arbor/article/viewArticle/1067
- Lysenko La teoría materialista de la
evolución en la URSS. Manuel Olarieta, 2012. Theoria. http://www.ucm.es/info/nomadas/trip/lysenko.html
- Todos los textos clásicos: Filosofía
zoológica de Lamarck, el origen de la especies y el Origen del hombre de Darwin, etc
23-¿Algo más que quieras añadir para finalizar la entrevista?
Solo agradecerte esta magnífica entrevista y animar a los lectores
a seguir explorando estas y otras ideas por su cuenta. También les invito a que
sigan las actividades y noticias de BiVOS a través de la web y de las redes sociales,
así como a que contacten con el equipo coordinador para cualquier duda interés
de participación.
[1]. He escrito “se le atribuye” con la constancia de que sus
precursores (Edward Blyth, William Wells, Patrick Mathew o Herbert Spencer) ya
habían explorado las mismas ideas con anterioridad, así como Alfred R. Wallace
quien publicó un artículo previo al libro de Darwin y llegó a las mismas
conclusiones de forma paralela. Del mismo modo, no he utilizado la palabra
evolución en mi introducción al pensamiento de Darwin del mismo modo que él
tampoco uso este término hasta la sexta edición del “Origen”.
Anexos:
-Artículo sobre el neo-darwinismo por Daniel Heredia, en este enlace.
-Video sobre el Manifiesto por una Biología del Siglo XXI aquí.